Шестиногие...
Низшие,..
Стрекозы,..
Тараканообразные
Прямокрылые...
Пухоеды,..
Равнокрылые...
Чешуекрылые
Сетчатокрылые
Жёсткокрылые
Перепончатокрылые
Двукрылые
"Микромиграции"
Вот что представляется мне наиболее важным и наименее систематически изученным: площадь среднего перемещения насекомых. В огромном большинстве случаев мы, например, не знаем, на какое расстояние - десятки или сотни метров - могут передвигаться при помощи ног насекомые, живущие на возделанном поле. Как же можно при таких условиях серьезно обсуждать вопрос о распределении популяций? Вот еще один источник ошибок в возможных расчетах! Некоторые ученые пытались определить величину перемещений с помощью радиоактивных изотопов: Джильес (1961) - для Anopheles gambiae, который перемещается меньше чем на полтора километра; Гудвин, Джэйнс и Дэвис (1957) - для Pissodes strobi, перелетающего на большие расстояния; Болдуин, Риордан и Смит (1958) - для Melanoplus, которые при обильном питании перемещаются всего метров на 30 почти за три недели, а на почве, лишенной растительности, на расстояние до 240 метров за 6 дней.
Роэр (1959) разработал интересный метод мечения гусениц капустницы, примешивая к их корму нейтральную красную краску. Краска передается взрослой особи, и таким образом можно проследить перемещения бабочек. Самцы остаются поблизости от места своего рождения, мигрируют только самки.
Бэйл, Элиасон и Илтс (1962) метили комаров, припудривая их мельчайшим порошком сернистого цинка, сильно флуоресцирующим, что позволило потом легко находить насекомых. Они также давали комарам Culex terralis подсахаренную воду с растворенным в ней производным родамина. Находили комаров по ярко окрашенным экскрементам. Но более любопытное наблюдение сделали Джойс и Роберте (1959): на не обработанном инсектицидами поле, если оно находится рядом с обработанным, повышается урожайность культур даже в том случае, когда исключена всякая возможность участия ветра, который мог бы разнести инсектициды. Следовательно, приходится допустить, что произошло перемещение вредителей к местам, оставшимся пустыми после применения инсектицидов. В описываемом случае речь идет о хлопковой плантации; авторы действительно установили, что если на поле, не подвергнутом дезинсекции, платность Empoasca и розовых червей не уменьшилась, то зато численность Hircothrips снизилась. Фауна данного поля лишь в том случае может быть совершенно независимой от фауны другого, предназначенного для обработки инсектицидами, если оно отстоит от него не менее чем на 150 метров. Не заложено ли здесь начало нового экологического опыта, который, быть может, позволит изучить во всех тонкостях проблему микромиграций - и активных и пассивных? Но надо сказать, что результаты эксперимента Джойса и Робертса на Hircothrips выглядят действительно очень странно: если предположить, что трипсы пассивно "дрейфовали" (в воздушных потоках), то почему бы им не дрейфовать дальше, к опустошенной зоне; если же предположить, что "дрейф" активен, то каким же образом трипсы узнали, что место освободилось?
В одной старой работе Флешнер (1950) пытается определить среднее расстояние, которое должен покрыть хищник, чтобы иметь шансы завершить свое развитие. Из трех видов личинок, поедающих Paratetranychus citri, божья коровка Stethorus picipes обследует за 1 час 15 минут площадь 312 квадратных сантиметров, Conventzia hazeni - 169, Chrysopa calijornica - 695. А жертва за то же время совершает странствие на 2320 квадратных сантиметрах. Оказывается, чтобы завершить цикл развития, для Stethorus достаточно одной жертвы на 6,5 квадратных сантиметрах, а для Chrysopa - на 169. Таковы лабораторные данные, которые следовало бы дополнить материалами, добытыми в поле. Это часть тех точных количественных данных, в которых мы испытываем острую нужду, хотя математики и считают задачу решенной.
Особенно важным кажется определение "средней площади странствий" для некоторых видов бабочек, которые, по-видимому, откладывают яйца на растении-хозяине совсем не с такой пунктуальностью, как было принято считать. По Детьеру (1959), личинкам очень часто бывает необходимо уметь вести активные поиски подходящего корма на полосе не менее 50 сантиметров; если они слишком удалены от пищи, смертность их может достигать 80% и даже больше, причем в выборе направления они способны руководствоваться лишь несовершенными показаниями своих чувств и только случайно попадают на растение - хозяина. Поэтому Детьер утверждает, что численность насекомых возрастает на возделываемом поле совсем не потому, что здесь больше растений-хозяев, а потому, что они здесь сближены.
Хенсон (1959) полагает, что можно выделить два типа насекомых: одни сосредоточиваются в одном и том же месте и в конце концов их популяции достигают там "невыносимых" плотностей, очень быстро истощая субстрат; у других же склонность к скоплению слабеет, когда плотность популяции чрезмерно возрастает, и они эксплуатируют среду, не слишком ее истощая (автор приводит интересный статистический метод, позволяющий вычислить "коэффициент скопления"). Измерение интенсивности группирования давно уже интересует экологов-математиков. Уотерс (1959) обсуждает применимые в этом случае статистические приемы. Не нужно думать, что в лаборатории или даже в банке с Tribolium насекомые не перемещаются, и об этом должны помнить те, кто определяет численность популяции, изымая для пробы часть муки. В банке с рисовыми отрубями, где разводят Ephestia саutella, все молодые гусеницы находятся в верхних слоях корма. Только начиная с третьего возраста они переползают в средние и нижние слои. Чем выше плотность, тем большее число особей в этих двух последних слоях. Гусеницы, готовые к окукливанию, наоборот, мигрируют в верхние слои (Такахаши, 1961). Некоторые ученые пытались экспериментальным путем вычислить коэффициент рассеяния. Нэйлор поселял Tribolium confusum на муке в круглых коробках, снабженных несплошными радиальными перегородками; это позволяло изучить степень скопления. Если не принимать в расчет пол насекомых, то можно считать, что тенденцию скопляться при низких плотностях популяции имеют имаго. При средней и высокой плотностях популяция распределяется равномерно. Но если учесть пол, то оказывается, что самцы более склонны собираться в группы, а самки - оставаться в изоляции. Автор смог установить, что запах сородичей, в особенности самцов, отталкивает самок, тогда как самцов запах сородичей, и в особенности самок, притягивает. Сакаи, Наризе и Хираицуми (1958) поместили плодовых мушек в систему банок, соединенных трубками. Сначала наблюдалась миграция, носящая случайный характер, затем, когда одна из банок оказывалась слишком перенаселенной,- массовая. Интенсивность миграции разных линий мушек варьировала в пределах 2-14%; в зависимости от линии менялась также и минимальная степень плотности популяции, которая служила для миграции толчком. Уже сейчас можно говорить о плодотворности этих опытов для генетики и экологии.
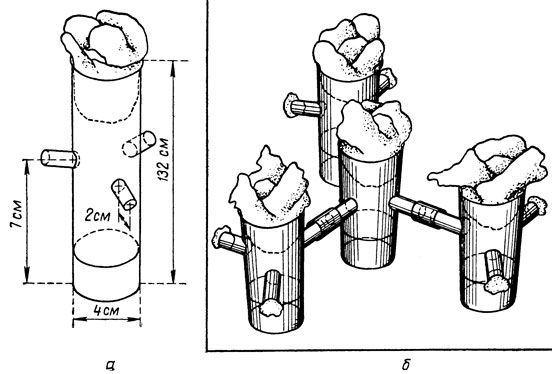
Установка для изучения микромиграций дрозофилы. а - трубка для выращивания одной популяции; б - четыре такие трубки, соединенные между собой (по Сакаи, Наризе, Хираицуми и Ияма)
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://invertebrates.geoman.ru/ 'Беспозвоночные (от простейших до насекомых)'