Шестиногие...
Низшие,..
Стрекозы,..
Тараканообразные
Прямокрылые...
Пухоеды,..
Равнокрылые...
Чешуекрылые
Сетчатокрылые
Жёсткокрылые
Перепончатокрылые
Двукрылые
Взаимодействие между паразитом и жертвой
Работ, посвященных этому вопросу как в практическом, так особенно и в теоретическом плане, настолько много, что здесь невозможно дать даже поверхностное представление о них. Ученые школы японского исследователя Утиды провели углубленные работы на перепончатокрылых из семейств птеромалид и браконид, паразитирующих на зерновке Callosobruchus chinensis. Когда особей всех трех видов помещали в один закрытый сосуд, численность обоих паразитов сначала повышалась за счет хозяина. При этом бракониды эффективнее сдерживали размножение зерновки при слабой плотности заселения, а птеромалиды - при повышенной; менялся и вес зерновок. Таким образом, "кормовая масса", которую они собой представляют для паразитов, не была постоянной. Выше всего был вес, когда в банке сидели одни зерновки или когда паразитов было немного; при большом числе паразитов вес зерновок заметно снижался. Впрочем, механизм колебаний веса (чрезвычайно нерегулярных) не совсем понятен. В присутствии обоих паразитов по мере повышения их плотности потомство хозяина явно убывало, однако влияние обоих паразитов было таким же, а не вдвое большим. Тем не менее Смит допускал, что действие нескольких паразитов более значительно, нежели одного; видимо, это не всегда так, по крайней мере в опытах Утиды.
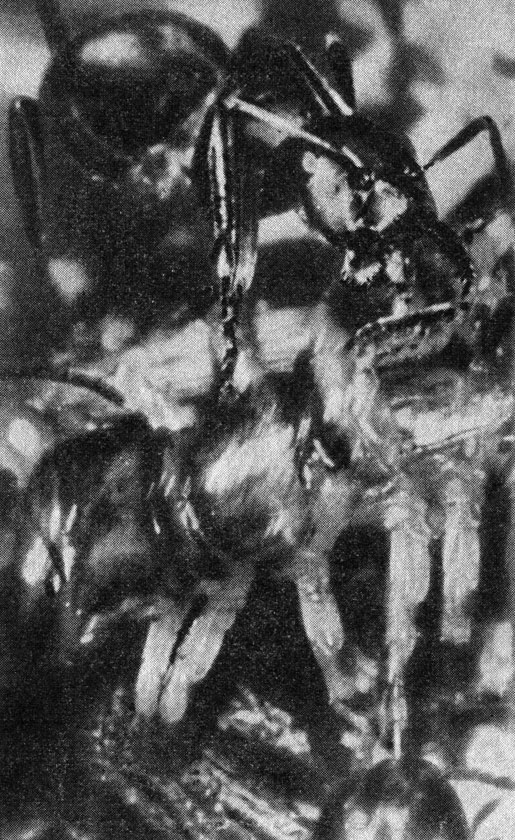
Муравьи, атакующие многоножку. Отношения между хищником и жертвой интересуют и эколога, изучающего, равновесие в животных сообществах, и агронома, желающего уничтожить вредных насекомых, не истребив при этом их естественных врагов
Проводя свое ставшее всемирно известным математическое изучение паразитизма, Никольсон постулировал существование регулярных колебаний численности обоих популяций, а также достижение точки равновесия по прошествии некоторого времени. В действительности, по крайней мере в рассматриваемом нами случае, колебания несомненны, но нерегулярны и резки; те, что касаются паразита, сдвинуты несинхронно с колебаниями в численности хозяина, да и установившаяся плотность или точка равновесия не так уж постоянна. Плотность культуры хозяина по прошествии известного срока склонна повышаться, а плотность культуры паразита падает, как если бы понижалась его биологическая жизнеспособность.
Наблюдаются даже некоторые парадоксальные явления. Например, если выращивать вместе Callosobruchus chinensis и С. quadrimaculatus, то через несколько поколений С. chinensis исчезнет. Зато введение одного паразитического перепончатокрылого сокращает одновременно популяции обоих видов зерновок, но они сохраняются в среде. Такахаши наблюдал не менее странное явление, выращивая амбарную огневку совместно с двумя различными паразитами (Habrobracon и Cimodus). Cimodus не оказывает очень уж серьезного влияния на численность популяции огневки, зато Habrobracon сильно сокращает ее, хотя огневке все же удается выжить; когда же на ней паразитируют оба вида одновременно, ее популяция сохраняет большую численность, чем при одном паразите.
Подобных случаев описано немало; на них построена целая математическая теория отношений между жертвой и ее врагами. Но, напоминаю, речь идет всегда о лабораторных исследованиях. А так ли уж много общего у паразитизма, наблюдаемого в ограниченном пространстве, на вредителе зерна с довольно специализированным питанием, с паразитизмом, наблюдающимся в природе? Конечно, общего немного... Отнюдь не желая обескураживать тех, кто занимается подобными исследованиями, я думаю, неплохо было бы посоветовать им шире варьировать условия экспериментов: что, скажем, произошло бы в сосудах, увеличенных в сто раз? Утида об этом подумал: когда он увеличил размеры сосуда для разведения насекомых, оставив неизменным количество корма, оказалось, что процент воспроизводства зерновок падает, а паразитов - повышается, и опять приходится сказать, что пока еще неизвестно почему.
Как полевые экологи критикуют математическую экологию
Я уже говорил выше, что не только постепенное развитие одновидовой популяции, но и взаимодействие паразита и его жертвы послужили мощным стимулом для математических исследований. Некоторые научные журналы почти целиком заполнены учеными выводами, представляющими всевозможные варианты анализа взаимоотношений между хозяином и паразитом.
Но реальность всегда богаче воображения, и приходится признать, что эти математические выкладки очень редко находят приложение в полевых условиях. Конечно, они не бесполезны, нет, но, повторяю, слишком еще много неизвестных нам параметров.
Как бы то ни было, нужно признать, что в проблеме "паразиты и их жертвы" математики неосторожно удалились от эксперимента. Томпсон, например, на основании своих вычислений допускал, что паразит должен полностью истребить своего хозяина, а следовательно, вместе с ним и самого себя. Но это значит забыть, что паразиты гораздо более чувствительны к климатическим изменениям, чем их хозяева, что доказано многими исследованиями. Одного только этого фактора более чем достаточно, чтобы не проявились предсказанные последствия "аккумулятивного паразитизма". Какая-нибудь экологическая катастрофа может привести к исчезновению паразитов, если они немногочисленны, или хозяев, если они очень сильно заражены этими паразитами, но расчеты показывают, что для этого потребуется значительно больший отрезок времени, чем тот, который предусматривается циклами Томпсона. Когда такие катастрофы следуют одна за другой через короткие промежутки, то они кончаются истреблением хозяина и его паразита; если их повторение происходит менее быстро, уничтожение хозяина паразитом замедляется, но не прекращается совсем. Чаще всего промежуток от катастрофы до катастрофы таков, что между хозяином и паразитом устанавливается некое "неустойчивое равновесие", при котором поддерживается существование обоих.
Еще один недостаток используемых математиками-экологами методов состоит в том, что параметры они выбирают произвольно, а аксиомы - неудачно. Вот, например, их основные положения: когда нет ни препятствий, ни ограничений в пространстве, корме, условиях среды, популяция возрастает в геометрической прогрессии; численность популяции возрастает непрерывно, и этот процесс можно понимать как сложение бесконечно малых величин, к которым применимо дифференциальное исчисление; плотность популяции в каждую единицу времени определяет ее возможное дальнейшее увеличение или его задержку.
На этих положениях основывается ряд других, относящихся уже к популяции паразита. Их главная мысль - популяция паразита будет действовать на популяцию хозяина, которая ведет себя согласно трем вышеизложенным правилам.
Но, как замечает Уллиетт, правила эти не точны. Во-первых, возрастание популяции в геометрической прогрессии осуществляется лишь в том случае, если его не прерывают сезонные или физиологические факторы. Но так не бывает, пожалуй, ни в одной среде, разве что в банке с мучным хрущаком или дрозофилой (да и то - как знать!..), а уж в природе - никогда. Даже в лаборатории - и это известно каждому наблюдателю - у животных обнаруживаются, и очень часто, большие колебания способности к воспроизводству при всех прочих равных условиях, что до сих пор невозможно объяснить. Так что истинная кривая популяции - это ряд пиков, разделенных глубокими впадинами; она почти совсем не поддается анализу с помощью бесконечно малых величин. Математик выходит из положения, соединяя вершины непрерывной линией, беря образцы через определенные равные промежутки или рассматривая только популяцию взрослых особей. Впрочем, все эти приемы можно считать законными при одном условии: помнить, что этими методами пользовались.
Перенаселенность, которая приводит к самоограничению популяции, сама зависит от ряда факторов, совсем не обязательно связанных только с количеством корма. Блестящий пример тому мы уже видели в исследовании Ле Ге Бреретона. В любом случае последствия перенаселенности очень сложны, трудны для истолкования и меняются от стадии к стадии. Например, у зеленых падальных мух, которые развиваются на трупах, молодые стадии в гораздо большей степени, чем взрослые, чувствительны к внутривидовой и межвидовой борьбе, но от паразитов больше страдают куколки.
Не нужно забывать, что среда не остается неизменной, она, как говорят американцы, кондиционирована присутствием живых существ. И это верно не только для Tribolium, но и для падальной мухи, личинки которой выделяют энзимы, вызывающие переваривание пищи вне организма. Даже тли сильно влияют па растение, которое сосут: выделяемые тлями вещества приводят к скрытым изменениям в обмене веществ растения или уродливому разрастанию тканей (галлам).
Уллиетт замечает также, что одна и та же плотность популяции может быть следствием самых различных факторов. А следовательно, ее нельзя рассматривать изолированно.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://invertebrates.geoman.ru/ 'Беспозвоночные (от простейших до насекомых)'